อากาศเปลี่ยนแปลง
ย่อมมีผลกระทบต่อพืช
Pollution spots ("weather fleck")
or physiological spots.
In fact, the symptoms of "weather fleck" are induced by ozone (O3) which is an air pollutant affecting many plants. This gas is produced secondarily in the atmosphere as a result of chemical reactions involving primary pollutants such as nitrogen dioxide (NO2) and hydrocarbons (HC), this under the influence of solar radiation.

it is considered the most important air pollutant. It enters the leaves through the stomata and affects mesophyll cells. It causes many small spots, located mostly on the upper side of lamina, sometimes contiguous and located along the midrib and secondary veins . These spots are often found on basal leaves. Their colour changes over time and turns fairly quickly from brown at first, to greyish white afterwards . When the attacks are very severe, whole leaves are affected. They age prematurely and fall.
Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants

Ozone Stress
Ozone is among the major components of photochemical air pollution responsible for causing significant damage to both cultivated plants and forest trees (Wang et al., 2007). Ozone enters to mesophyll cells via stomata where it immediately interacts with water and other cellular components to generate phytotoxicity mainly by elevating the generation of ROS (such as , H2O2,.OH, and 1O2), and triggering a series of signaling cascades and plant defense responses (Long and Naidu, 2002; Ashmore, 2005). Extensive reports are available on the key regulatory roles of SA in plant-ozone stress tolerances (Yalpani et al., 1994; Sharma et al., 1996; Yoshida et al., 2009; Khan et al., 2012a; Pál et al., 2014). SA can work as a signal molecule and promote molecular and physiological changes in ozone-exposed plants (Tamaoki, 2008). Moreover, SA is required to potentiate the antioxidant response, maintain cellular redox state, and activate processes against hypersensitive cell death and ozone-sensitivity (Rao and Davis, 1999). SA was reported to maintain plant growth, development and cellular redox system by activating the GSH-biosynthetic pathway (Yoshida et al., 2009).

In ozone-exposed A. thaliana, SA was involved in the accumulation of defense-related transcripts and induced resistance (Sharma et al., 1996). SA is also involved in signaling network integrating other phytohormones such as JA and/or ethylene in ozone-exposed plants (Rao et al., 2002). Notably, both SA and ethylene were evidenced by these authors to act in concert to regulate ozone-induced cell death in A. thaliana. In the same plant, ozone-mediated induced biosynthesis of JA or methyl jasmonate was reported to attenuate SA-dependent lesion-initiation that eventually was considered as a major factor for the decreased lesions caused by ozone (Rao et al., 2000). It has also been evidenced that ozone-accrued SA-accumulation can be promoted by ethylene-mediated regulation of the expression of the PAL and chorismate mutase (CM) genes in ozone-exposed Nicotiana tabacum (Ogawa et al., 2005). In another study, ozone-sensitivity in hybrid poplar was correlated with insensitivity to both SA and jasmonic acid, where these phytohormones were associated with the programmed cell death in lesion formation (Koch et al., 2000).

Temperature Stress
In the current changing environmental scenario, both low (cold and chilling stress) and high (heat) temperatures have become a potential abiotic stress-threat to crop plants. Temperature stress affects many plant-physiological and biochemical processes and induces molecular mechanisms and gene expression to modulate plants responses (Larkindale and Knight, 2002; Khan et al., 2013a,b; Kazemi-Shahandashti et al., 2014; Siboza et al., 2014). SA-supplementation has been reported to differentially benefit several plant species exposed to low/chilling temperatures (Janda et al., 1999; Ding et al., 2002; Horváth et al., 2002; Kang et al., 2012; Kazemi-Shahandashti et al., 2014; Siboza et al., 2014) and high (He et al., 2002; Larkindale and Knight, 2002; Clarke et al., 2004; Shi et al., 2006; Wang and Li, 2006; Wang et al., 2010; Khan et al., 2013a,b). SA (0.5 mM) modulated antioxidant enzymes (such as ascorbate peroxidase, APX; superoxide dismutase, SOD; guaiacol peroxidase, GPOX; GSH reductase, GR) and improved chlorophyll fluorescence in Z. mays under low (2°C) temperature stress (Janda et al., 1999). Exogenously SA can inhibit the activity of isozymes (CAT-1 and CAT-2) of catalase (CAT) which in turn can mediate responses of Z. mays to low temperature stress (Horváth et al., 2002).

Salicylic acid (2.0 mM)-mediated increased synthesis of total phenolics and the activity of PAL were reported to improve chilling tolerance in cold-stored lemon fruit (Citrus limon; Siboza et al., 2014). Mutlu et al. (2013) reported that exogenously sourced SA results in cold tolerance by enhancing antioxidant enzymes, ice nucleation activity, and the patterns of apoplastic proteins in H. vulgare genotypes. In another report, SA significantly protected ultra-structures in Musa acuminata seedlings under chilling stress (Kang et al., 2012). Least reports are available in literature on the molecular mechanisms underlying SA-mediated improved plant tolerance to cold/chilling temperature. Increased chilling tolerance was evidenced in chilling-exposed S. lycopersicum fruit as a result of low concentrations of (0.01 mM) MeSA-mediated induction in the synthesis of some stress proteins, such as PR proteins (Ding et al., 2002). The applied MeSA significantly increased accumulation of PR-2b and PR-3amRNAs but slightly increased PR-3b mRNA accumulation (Ding et al., 2002).
Salicylic acid-mediated improved plant tolerance to heat stress has also been reported (He et al., 2002; Larkindale et al., 2005; Wang et al., 2010; Khan et al., 2013a,b). Larkindale and Knight (2002)reported that the transgenic Arabidopsisseedlings showing a bacterial SA-decomposing salicylate hydroxylase were less tolerant to heat stress. SA signaling played an important role in the acquisition of Ocimum basilicum to heat (Clarke et al., 2004). SA application increased antioxidant system in Vitis vinifera and induced activity of APX, GR, and monodehydro ascorbate (MDHA), increased redox ratios of AsA and GSH, and maintained Ca2+-homeostasis were reported in SA-supplemented and cold/heat treated V. vinifera (Wang and Li, 2006). SA (1.0 mM) decreased electrolyte leakage and oxidative stress, and improved maximum yield of PSII, Fv/Fm, and the quantum yield of the PSII electron transport in Cucumis sativa seedlings after both heat stress and recovery (Shi et al., 2006). Recently, Khan et al. (2013b) have shown that treatment of 0.5 mM SA can alleviate heat stress in T. aestivum by increasing Pro-production and restriction of the stress ethylene formation under heat stress. Notably, the details of the mechanisms of heat stress mitigation with the application of SA are not available and the area could be an open challenge at both physiological and molecular levels.
M. Iqbal R. Khan,1 Mehar Fatma,1 Tasir S. Per,1 Naser A. Anjum,2,* and Nafees A. Khan1

Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants

M. Iqbal R. Khan,1 Mehar Fatma,1 Tasir S. Per,1 Naser A. Anjum,2,* and Nafees A. Khan1
Abstract
Abiotic stresses (such as metals/metalloids, salinity, ozone, UV-B radiation, extreme temperatures, and drought) are among the most challenging threats to agricultural system and economic yield of crop plants. These stresses (in isolation and/or combination) induce numerous adverse effects in plants, impair biochemical/physiological and molecular processes, and eventually cause severe reductions in plant growth, development and overall productivity. Phytohormones have been recognized as a strong tool for sustainably alleviating adverse effects of abiotic stresses in crop plants. In particular, the significance of salicylic acid (SA) has been increasingly recognized in improved plant abiotic stress-tolerance via SA-mediated control of major plant-metabolic processes. However, the basic biochemical/physiological and molecular mechanisms that potentially underpin SA-induced plant-tolerance to major abiotic stresses remain least discussed. Based on recent reports, this paper: (a) overviews historical background and biosynthesis of SA under both optimal and stressful environments in plants; (b) critically appraises the role of SA in plants exposed to major abiotic stresses; (c) cross-talks potential mechanisms potentially governing SA-induced plant abiotic stress-tolerance; and finally (d) briefly highlights major aspects so far unexplored in the current context.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4485163/

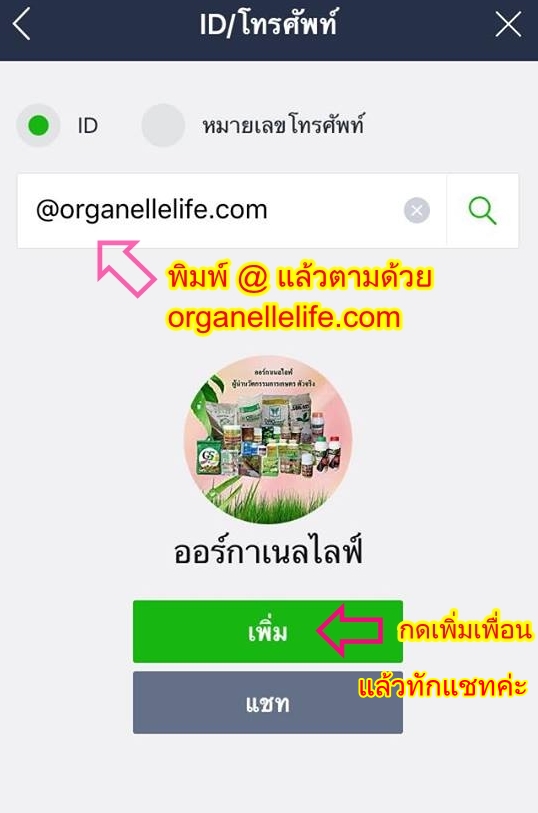
ปรึกษาเรา
หรือกดลิงก์ด้านล่าง แล้วเพิ่มเป็นเพื่อน เพื่อคุยสอบถามข้อมูลได้ครับ http://line.me/ti/p/%40organellelife.com
ไม่มีความคิดเห็น:
แสดงความคิดเห็น